|
|
|
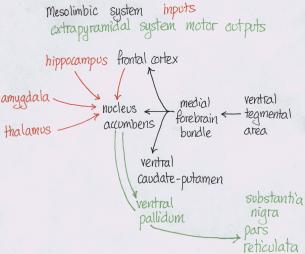
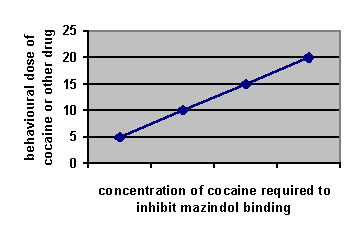
Amphetamine and cocaineAddiction is when the drug rules your life; everything is geared round the drug. Dependence is when you need the drug to perform normally, e.g. cigarettes where you don’t get a high. Abuse is when it causes problems, e.g. with family or work. Dopamine is made in the cytoplasm. The precursor of dopamine, and therefore the other catecholamines, is tyrosine. The limiting factor on tyrosine conversion into dopa is tyrosine hydroxylase. AMPT competes with tyrosine hydroxylase for its binding sites and therefore is a tyrosine hydroxylase inhibitor. From there it’s put into vesicles, which are later released into the synapse given an action potential. Spike-dependent release is dependent on there being Ca2+ ions extracellularly, and reserpine, which blocks vesicular uptake of dopamine, will stop it. Dopamine is also released continuously from the cytoplasm in nerve endings into the synapse in a spike-independent manner. Spike independent release is affected by AMPT. Amphetamine and cocaine 1) decrease dopamine synthesis, 2) increase release of dopamine into the synapse, 3) block dopamine reuptake back into the cell, and 4) inhibit monoamine oxidase, the enzyme that breaks catecholamines down. Cocaine has most effect on inhibiting reuptake, while amphetamine has most effect in increasing synthesis of dopamine. Amphetamine increases the spike-independent spontaneous release because in vivo and in vitro microdialysis studies show that dopamine release in the presence of amphetamine is reserpine and Ca2+ independent and AMPT dependent. An action potential will still release dopamine in the normal way. Cocaine increases the spike-dependent release of dopamine from vesicles because dopamine release is Ca2+ and reserpine dependent and AMPT independent. Cocaine acts by plugging the transporter on the presynaptic membrane that would normally transport released dopamine back into the presynaptic neuron. There are four methods for investigating reward: 1) intracranial self-stimulation, 2) drug self-administration, 3) conditioned place preference, and 4) drug discrimination. intracranial self-stimulationFrom intracranial self-stimulation, where rats press levers to pass a small current through electrodes in particular areas of the brain, we see that it’s the mesolimbic dopaminergic system that is important. It seems to be the innervation of the nucleus accumbens that is important in the reinforcing properties of psychostimulants. Positive intracranial self-stimulation sites are confined to dopaminergic areas in the mesencephalon, including the nucleus accumbens (overactive in schizophrenia), the striatum (underactive in Parkinson’s) and areas of the prefrontal cortex. The hottest area is in the medial forebrain bundle, which if stimulated will activate the ventral tegmental area. Intracranial self-stimulation also occurs for stimulation of the ventral tegmental area, which causes increased dopamine release in the medial forebrain bundle, as shown by increased dopa/dopamine and HVA/dopamine levels through microdialysis. There’s also increased dopamine in the striatum. The substantia nigra doesn’t mediate intracranial self-stimulation.
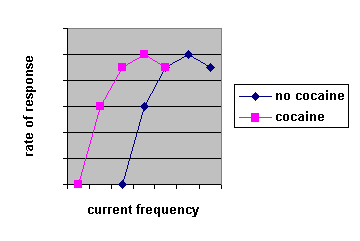
You can’t just look at what administering a drug does to intracranial self-stimulation rates of lever pressing because the drug will likely also have motor effects. Two techniques used are the set-reset technique and curve shifting. In the set-reset technique a rat presses a lever for intra-cranial self-stimulation, but after every say 5 presses the frequency or intensity goes down. They can at any point go to another area of the room and press another lever to put the current back up to its original value. If amphetamine is administered they have a lower reset threshold, implying psychostimulants increase the rewarding properties of intracranial electrical stimulation. In the curve-shifting technique, either frequency or intensity is held constant and the other is increased. as your independent variable and rate of bar pressing is measured as your dependent variable. What you find is that as you increase your current frequency or intensity bar pressing goes up to some maximum value then stays constant. If you administer psychostimulants the curve is shifted to the left; i.e. they’ll bar-press more for the same current. Again, this implies psychostimulants increase the reinforcing properties of reward.
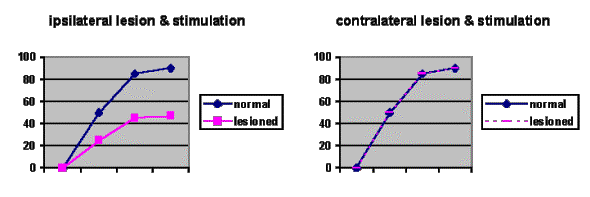
drug self-administrationFrom drug-self administration studies we can confirm that it is the mesolimbic dopaminergic system that is important. Surgery is performed putting a catheter into the animal’s jugular, which is then linked to a computer and lever so if the animal bar-presses it gets a dose. They’re given a priming dose to get them going. You see animals will readily self-administer for amphetamine or cocaine. If the nucleus accumbens or ventral tegmental area is lesioned using 6-hydroxy dopamine, extinction occurs, but not if the striatum or frontal cortex is lesioned.
Another way to look at this is to look at breakpoints. If you increase the ratio reinforcement schedule progressively, eventually the animal will stop responding. If you give them a D2 antagonist, breakpoints increase. conditioned place preferenceYou can confirm the same findings using conditioned place preference. In this paradigm an animal can move freely between 2 rooms, each with distinctive colours or textures. The drug is administered in one of the rooms; say drug on one side one day, saline on the other the next, etc. The animal will associate the neutral stimuli of the environment with the drug, and spend more time there if it’s rewarding. In a biased design the animal is conditioned to whichever of the two rooms it initially spent less time in. if you stop administering amphetamine or cocaine the behaviour will eventually be extinguished. If you lesion the nucleus accumbens (or use a D2 antagonist like haloperidol), conditioned place preference for amphetamine will be extinguished, but not for cocaine. Cocaine is not a pure dopamine agonist. It also has serotonergic effect and affect in other areas. If you put a tiny amount of amphetamine directly in the nucleus accumbens you still get conditioned place preference. drug discriminationDrug discrimination studies further confirm that it is action at the dopamine receptors that is rewarding. This is not a measure of reward, but similarity or dissimilarity. An animal is given either the drug or saline, with the drug being the discriminatory stimulus for some other operant behaviour, say bar pressing for food. Rats will generalise from cocaine to other dopamine agonists or reuptake inhibitors. The administration of a D2 antagonist will disrupt their ability to distinguish between cocaine and saline, and a D1 antagonist will partially disrupt it.
|